What is the structure and physiology of Crustacea? Information about appendages, cuticle, nervous system, digestive system etc. of Crustacea.
Source: pixabay.com
To allow movement, the rigid crustacean shell is broken into a series of segemental rings, or somites, connected by flexible membranes. The earliest crustacean probably had a large and indefinite number of somites; some existing crustaceans have more than 60, but most have between 16 (Copepoda) and 21 (Malacostraca). The most anterior part of the head (the acron) and the most posterior part of the abdomen (the telson), which bears the anus at its base, are not considered true somites.
Appendages.
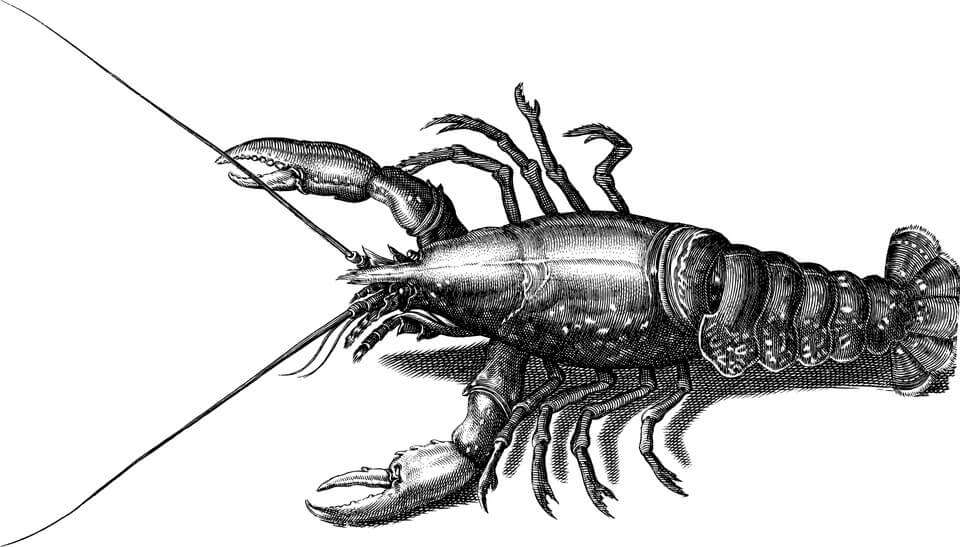
Typically the crustacean body is divided into three major regions: head, thorax, and abdomen. In some groups the head is fused with some of the thoracic segments to form the combined structure known as the cephalothorax. Each body region bears a number of appendages, and each appendage consists of a number of joints, or segments. The stem, or base, segment, known as the protopodite, is composed of two parts: the coxopodite, which is joined to the body, and the basipodite. Characteristically, the appendages have two branches, attached to the basipodite. The inner branch, arising closer to the middle of the body, is called the endopodite; the outer branch is called the exopodite. Both the endopodite and the exopodite may themselves be segmented.
One of the two branches may be lacking on certain appendages in some crustaceans. The pincers (chelipeds) and walking legs (pereiopods) of the lobster, for example, are formed from only the stem (protopodite) and endopodite, the exopodite branch being absent.
The five pairs of head appendages are the first antennae (or antennules), the second antennae, the mandibles, the first maxillae (or maxillulae), and the second maxillae (or maxillae). The first antennae are typically un-branched, although in many Malacostraca they have two branches. Both pairs of antennae are primarily sensory, but they may have other functions, most commonly locomotion or feeding. The mandibles serve to chew the food, but the outer-end segments (palps) may aid in locomotion or in setting up feeding currents. Both pairs of maxillae, which tend to be broad, assist in feeding.
The thoracic appendages serve a great variety of functions in different crustaceans. One to three of the first three pairs may be modified to assist in feeding, in which case they are called maxillipeds. Other appendages may be adapted for grasping or for locomotion. Often the posterior thoracic limbs or the anterior abdominal limbs are modified to serve in reproduction. Abdominal limbs are absent in many groups of crustaceans. When present, they are commonly used for setting up currents of water, for swimming, or for carrying eggs and young.
Cuticle.
The crustacean exoskeleton, or cuticle, is secreted over the whole body surface by the epidermis, a single layer of cells covering the body. The cuticle is composed mainly of protein and chitin and varies from a thin, flexible sac to a thick, rigid shell. It is usually stiffened and hardened by the deposition of calcium salts within it and by a tanning process in which the protein part of the cuticle is hardened by qui-nones derived from the blood. The cuticle often sends projections and folds into the interior of the body; these serve as attachment points for muscles and as braces for the body. Externally the cuticle forms a wide variety of spines and hollow projections, called setae, which have many different functions.
Structurally the cuticle consists of four layers: a thin, outer, nonchitinous epicuticle and three chitinous layers of endocuticle—the pigmented layer, the calcified layer, and the uncalcified layer. The epicuticle and pigmented layers, as well as the calcified layer, are calcified.
Eyes.
Two types of eyes occur in crustaceans: simple eyes, located centrally (median eyes), and compound eyes, located on either side. Median eyes occur in the nauplius stage of the larvae and are the only eyes in copepods, adult barnacles, and most ostracods. Such an eye consists of two or more sensory cells surrounded by a cup of screening and reflecting pigments. Sometimes a lens is present. The compound eye is similar in structure to that of insects, consisting of a number of tubular units, called ommatidia, radiating out from the end of the optic nerve. The outer surface of each omma-tidium is covered by a corneal lens, a transparent bit of cuticle secreted by two underlying epidermal cells. Below the epidermal cells is the cylindrical or conical crystalline cone, a lenslike structure leading to the retinula. The retinula, consisting of light-sensitive nerve cells, surrounds the translucent, light-refracting rhabdome, or optic rod. Nerve fibers from the retinula enter the optic nerve.
Each ommatidium is isolated from the others by two groups of pigment cells, one group around the crystalline cone, the other around the retinula. The pigment moves within the cells to cover more of the ommatidium in bright light, less of it in dim light.
Hearing.
No special hearing organ, or ear, has been found in the Crustacea, but numerous hollow setae, supplied with nerves, are present on the surface of the body and the appendages, and move when touched. It is possible, therefore, that these and other body parts can respond to and detect vibrations, including sound.
Smelling and Taste.
Because the detection by aquatic animals of dissolved substances in the water around them cannot readily be separated into smelling and tasting, it is best to refer to this sense as chemoreception. Chemoreception is extremely important to crustaceans, and they are very sensitive to chemical stimuli. These stimuli are detected by a variety of structures, especially the thin-walled hairs found in greatest numbers on the antennae.
Statocysts.
Organs of equilibrium, the stato-cysts, occur in the Malacostraca, usually in the basal segment of the first antenna, but sometimes in the tail fan (uropods and telson). A statocyst consists of a rounded, fluid-filled sac lined with sensory cells that often have hairlike projections arising from their surfaces. One or more stonelike statoliths lie inside the sac in contact with the hairs. As the body position changes, the statoliths press on different groups of hairs, and the crustacean is aware of its position in relation to the pull of gravity. Other hairs may be affected by movement of the statocyst fluid and give the crustacean information about the rate of turning.
Nervous System.
As in other arthropods, the nervous system is basically of the “ladder” type, consisting primitively of a pair of ventral (“bottom”) nerve cords with a cross-connecting ganglion (mass of nervous tissue) in each somite. In all but the more primitive Crustacea, however, the two cords are fused.
The brain is composed of the fused ganglia of the first and second antennae and the eyes. Nerve cords from the brain circle down past the esophagus to join the subesophageal ganglion, composed of the fused ganglia of the mouthparts. Varying degrees of fusion of the ventral ganglia have developed independently within most crustacean groups. In most crabs all ventral ganglia are fused with the subesophageal ganglion to form a single mass.
Respiration.
In very small crustaceans, respiration takes place through the general body surface, especially in certain specialized areas of the integument (outer covering). Integumentary respiration may also occur to some extent in crustaceans that rely mainly on other methods.
Gills are the usual respiratory organs, however, and they are typically associated with the appendages. The typical gill in the largest group of crustaceans (the Malacostraca) is a thin-walled structure, well supplied with blood vessels, that arise from the base of a thoracic limb.
Adaptations for aerial respiration occurring in terrestrial and semiterrestrial crustaceans comprise a progressive reduction in the number and size of the gills. Air provides more available oxygen than does water, and so less gill area is needed for aerial respiration. Reduction of the gills also diminishes the amount of body water lost through the gills. Land crabs, with their relatively higher metabolic rate and greater need fcyr oxygen, compensate for gill reduction by changes in the blood to increase its affinity for oxygen and by enrichment of the walls of the gill chamber with blood vessels to facilitate the exchange of oxygen and carbon dioxide.
Digestive System.
The crustacean digestive system is a relatively straight tube consisting of three parts: foregut, midgut, and hindgut. The foregut and hindgut are lined with a chitinous layer, which is shed and replaced at each molt. The foregut may be only a tubular esophagus, but in most malacostracans it widens to form the cardiac part of the stomach, the walls of which bear grinding ridges, teeth, and calcareous ossicles, moved by muscles. The cardiac stomach chews the food, and its teeth are reduced or absent in decapods, whose mandibles have sharp teeth for chewing food. Tubular digestive structures called cecae, open into the midgut; in higher crustaceans one pair of cecae has developed into a large digestive and absorptive organ known as the hepatopancreas.
Excretion.
Paired excretory organs open on or near the bases of the second antennae or the second maxillae. Both antennal and maxillary glands are commonly present in larvae, but usually only one pair persists in the adult. The principal nitrogenous waste product is ammonia, but urea, uric acid, and amines are also excreted. A significant amount of the wastes may be excreted by diffusion through the epithelium of the gills.
Circulatory System.
The vascular, or circulatory system is quite variable. In its most complete form, in the Malacostraca, the system includes a heart, accessory hearts, arteries, capillaries, and venous sinuses. The heart or the arteries or both may be absent in some lower crustaceans, the blood being propelled by body movements.
mavi
