What are the characteristics of parasites? Information about parasites relationships to hosts, parasitic organisms, effects on hosts.
PARASITES, are organisms that live in or on other organisms (called hosts), at whose expense they obtain food, shelter, or some other benefit. The parasite-host association resembles the predatoryprey association in that both parasites and predators benefit from the association, while both hosts and prey are harmed by it. Parasites generally differ from predators in several respects, however. Parasites are typically smaller than their hosts, while predators are typically larger than their prey.
Also, parasites are typically adapted to live in or on one kind of host, while predators usually are not highly specialized in their prey. Unlike predators, parasites generally do not kill their victims outright, and many hosts live out their normal life span while supporting a population of parasites. In fact, parasite-host associations that have evolved over a long period of time tend to become harmless or even beneficial to the hosts. Certain intestinal bacteria, for example, that originally may have been harmful to their hosts, now help the hosts digest their food—parasitism has thus evolved into mutualism (symbiosis).
The term parasite is sometimes extended to apply to organisms that become a burden to other organisms because of their social relationships. For example, cuckoos and cowbirds lay their eggs in the nests of other birds, which then are saddled with the burden of rearing the alien young—often at the expense of their own. Colonies of certain ants live in the nests of other ants, which provide them with food and shelter. And some human beings live at the expense of their fellow men without contributing anything in return.
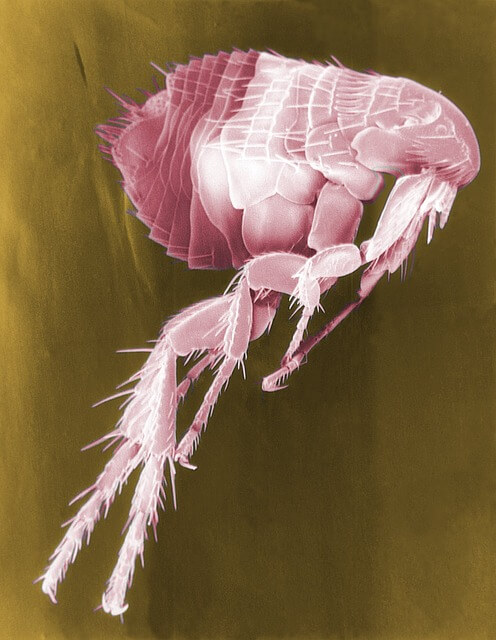
Flea
The parasitic mode of life undoubtedly originated soon after there were hosts to parasitize and has been resorted to by members of almost every major group of plants and animals and all groups of microorganisms. The principal groups of parasitic organisms are briefly considered below. Most kinds of plants and animals, even many microorganisms, serve as hosts, and most of them harbor a number of kinds of parasites.
There are actually many more parasites than hosts in the world, though fewer species of them, because parasites, living in environments that are relatively more stable than those of their hosts, undergo evolutionary changes more slowly. Whole groups of hosts, now differentiated into separate genera, families, or even orders, and in some cases now separated by oceans or continents, may harbor identical or nearly identical parasites merely because these inhabited the remote ancestors of the group and remained with them through the ages. The ostrich and the rhea, for instance, though one now lives in Africa and the other in South America, harbor the same tapeworm. The parasites harbored by animals sometimes give better clues than do their anatomical characters to their evolutionary affinities and past distribution.
Relationships to Hosts.
Parasites, like commensals, have been derived from free-living organisms ; by accident or experiment they have discovered the advantages of living at the expense of their fellow creatures, and have, in the course of evolution, adapted themselves to this life by more and more specialized modifications or changes in structure, physiology, and habits. Most parasites of animals originated as parasites of the skin or digestive tract or by inoculation by bloodsuckers.
The parasites harbored by a host are determined by satisfactoriness of the environment supplied by the host, which is sometimes very exacting, and by habits of the host which allow the parasites to gain access to it. Sometimes one of these criteria is of dominant importance, sometimes the other. There are some parasites that are so particular about their environment that they normally parasitize only a single species of host; for example, Taenia solium, the tapeworm, and Ancylostoma duodenale, the hookworm, of man.
In contrast the trichina worm, Trichinella spiralis, can develop in any meat-eating mammal, and the fluke, Heterophyes heterophyes, in practically any fish-eating mammal or bird. Some parasites are very particular about one host in the life cycle, but relatively indiscriminate about another. Certain strains of relapsing fever parasites, for instance, can develop in almost any mammal, but will tolerate only single species of the soft tick (Omithodorus), which serve as alternate hosts and transmitters.
Many parasites in the course of their life cycles make use of two to four different hosts in succession. The one in which sexual reproduction occurs is the definitive host, the others intermediate hosts. First intermediate hosts are sometimes ancestral hosts which were once defiftitive hosts; for example, the molluscan hosts of flukes and the crustacean hosts of many tapeworms.
Often, however, the intermediate host or hosts, and possibly all such hosts beyond the first one, have been adopted merely for convenience, to facilitate access to the final hosts. Usually such hosts are not very specific but there are exceptions. Man, strictly speaking, is an intermediate host, and a very specific one, for his malaria parasites, which reproduce sexually in mosquitoes of the genus Anopheles. On the other hand, man is the definitive host for the pork tapeworm, but quite a variety of mammals, including man himself, may serve as intermediate hosts.

Cayenne Tick
Parasitism is of all kinds and degrees. Some parasites are facultative, some obligatory; some live continuously on their hosts, others come and go; some are parasitic throughout their active lives or even generation after generation on one host, others only during certain portions of their life cycle, either as larvae (for example, botflies) or as adults (hookworms).
Parasites living on the surface of their hosts are called ectoparasites; those that live internally, endoparasites. There is, however, no sharp demarcation between these, since some living in such areas as the mouth, cloaca, and gills, or which partially or completely burrow into the skin, might be placed in either category. In vertebrate animals, parasites may be designated as intestinal, respiratory, and the like; or in the case of viruses, neurotropic, viscerotropic, and so on, according to the organs to which they are attracted. A distinction is also sometimes made between intestinal parasites, living in the lumen of the intestine, and parenteral ones, living in the tissues, that is, in the true interior of the body.
Effects of Parasitism.
Parasitism may have profound effects on parasites in the course of adaptation to their habitat and way of life. They are sometimes considered degenerate or degraded, but they are as beautifully adjusted to their environment as any organisms can be. They may combine extreme specializations, particularly in reproduction, life cycle, and methods for obtaining nourishment, with extreme reduction or loss of parts or functions no longer needed, such as organs of locomotion, special sense organs, and highly developed nervous system.
A tapeworm has no need for legs, fins, eyes or ears, or even an alimentary canal, and has retained no vestiges of them, but it can outstrip most animals in reproductive capacity and refinements in life cycle, and has developed a system of submicroscopic villi on its surface for absorption of food. The ultimate in parasitic adaptation is, perhaps, reached in the crustacean parasite of crabs, Sacculina. It starts out to be a barnacle but ends up as a shapeless mass of cells inside its host, from which food-absorbing roots spread throughout the body cavity, and from which a saclike structure grows to the outside of the host and, by hermaphroditic reproduction, becomes filled with eggs which are retained in the sac until they hatch as free-swimming larvae.
Physiological characters also become modified. Many parasites, living on blood or sap or in the tissues of their hosts, lose many of the metabolic functions which are necessary for their free-living relatives. Many free-living bacteria, for instance, can synthesize everything they need from simple inorganic chemicals; for parasites much of this may be superfluous, and these often come to depend on the host for vitamins and other metabolic needs.
In parasites that live inside the cells of their hosts even the enzymes used for respiration, synthesis, and the like, may be lost and those of the host cell utilized. This process reaches its ultimate in some of the plant viruses which have shed all their own enzymes and chemical accessories and are reduced to naked nucleoprotein molecules. Some parasites, on the other hand, have special physiological equipment to enable them to take blood or prevent its coagulation, penetrate tissues, resist antibody action.
Transmission.
The most remarkable specializations of parasites are their amazing reproductive capacities and the almost incredible modifications in life cycle to provide safe conduct for them from host to host, and thus insure the survival of the species when the host dies. .Every parasite not only must have a means of access for itself but also some means of exit for its offspring, even though in a few instances this is only accomplished by the devouring of the host by a predator.
The means of access may be onto or through the surface, either by body contact (lice), active migration (fleas), wind dispersion (molds), burrowing (hookworms), passive transfer by flies (trachoma virus), or inoculation by vectors such as aphids, mites, mosquitoes, and leeches. Vectors may be simple mechanical carriers, such as houseflies or roaches; they may serve as culture tubes in which the parasites multiply, for example, fleas for plague; they may be invaded by larval organisms and then serve merely for transport of them; or they may serve as true intermediate hosts in which a part of the life cycle must be passed, either with accompanying multiplication (for example, malaria parasites in mosquitoes), or without multiplication (for example, filarial worms in mosquitoes or guinea worms in Cyclops).

Parasite
In the case of animals, transmission may also be via the respiratory system, for instance, influenza virus ; or, more commonly, the mouth and alimentary canal. This transmission via the mouth may take place by accidental contamination of food or water by bacteria, viruses, cysts, and eggs; by licking eggs from the skin (bots) ; or by eating intermediate hosts harboring certain Protozoa or helminth larvae. When ancestral intermediate hosts do not serve as food for the definitive host (for example, snail hosts of bat flukes), second or even third intermediate hosts which are eaten may be involved (for example, for the bat flukes, immature aquatic insects which will eventually become aerial). One fluke, Alaria mustelae, parasitizes successively a snail, a tadpole, and a frog or mouse before reaching its definitive host, a weasel.
Exit for parasites is most frequently through the surface or with the excreta, in the case of metazoan parasites either as eggs or larvae. Vectors may again be involved for the release of blood- or skin-dwelling parasites as, for example, trypanosomes, malaria parasites, and filariae.
Effects on Hosts.
Parasites affect their hosts by mechanical injury, such as devouring blood or tissues, blocking ducts or vessels, causing bleeding, and providing portals of entry for smaller organisms; by robbing them of vitamins or other essential substances; and by toxic effects, either by liberating toxic products or by stimulating allergic reactions. Often several types of injury occur together. In most cases it is to the advantage of the parasite as well as the host that the injury to the latter be minimal, and in host-parasite relations of long standing usually some degree of mutual tolerance develops—a sort of armed truce.
A host may protect itself by increasing resistance to harmful effects either in the individual or, by natural selection, in the species or race. Higher animals may increase resistance by the development of antibodies either against the parasite itself, against its toxic products, or against the enzymes which enable it to grow, multiply, or defend itself against the host. Anything that decreases the host’s resistance, for example, poor diet or exhaustion, may permit the parasite to become injurious again, whereas improved resistance may lead to complete elimination of the parasites.
Capacity for resistance often depends on genetic characteristics, hence the differences that frequently exist between races or strains of the same plants or animals in their reaction to parasites. However, parasites too are capable of individual and genetic changes, and may develop increased resistance not only to the host’s defense mechanisms, but also to drugs or chemicals used against them. Acquired drug resistance of parasites is one of the greatest obstacles to successful chemotherapeutic treatment and to control of flies and other vectors by chemicals.
Diagnosis, Treatment, and Control.
Diagnosis of parasitic infections is sometimes possible by means of symptoms, especially in plants; but in animals it is often necessary to resort to examination for the parasites or their eggs or larvae, to culture methods, or to biochemical tests or immunological reactions, including skin tests.
Treatment involves either removal or destruction of the parasites by such means as drugs, chemicals, X-rays, or surgery; interference with their metabolism or welfare by use of antibiotics or other drugs, or by injection of immune or immunizing serums; or by such supportive measures as repair of injuries and enhancement of resistance by improved diet, rest, and the like. Often the supportive treatment is quicker and longer lasting than elimination of parasites which may be quickly reacquired; sometimes the anti-parasite treatment cannot be tolerated until the patient’s health has been improved. Then, as often as not, the host eliminates the parasites without further outside help.
To control parasites effectively, full knowledge of their habits and life cycles is needed so that the weakest link or links in the chain of events necessary for their survival or dispersion can be attacked. This may involve destruction of the parasites in or on their plant or animal hosts ; destruction of spores, cysts, eggs, or nonparasitic stages outside the host, in soil, water, or food; the elimination of intermediate, alternate, or reservoir hosts (for example, snails for schistosomes, barberry for mold parasites of wheat, or rats for plague) ; breaking the mechanism by which the parasites are transferred from the environment or from another host; or protection of the host against invasion of the parasite by such means as mechanical protection of the skin, incorporation of drugs in the water or food of animals, or vaccination; and sometimes selective breeding of resistant stock.
Parasitic Organisms.
Although parasitism has developed in almost every major group of organisms, including even vertebrate animals (lamprey and hagfish) and flowering plants (mistletoe), it is especially prevalent among microorganisms (viruses, bacteria, fungi, and Protozoa) and certain groups of invertebrates, the major ones of which are various helminths and arthropods. Parasitism has arisen to a lesser degree among mollusks (larvae of freshwater mussels) and annelids (leeches).
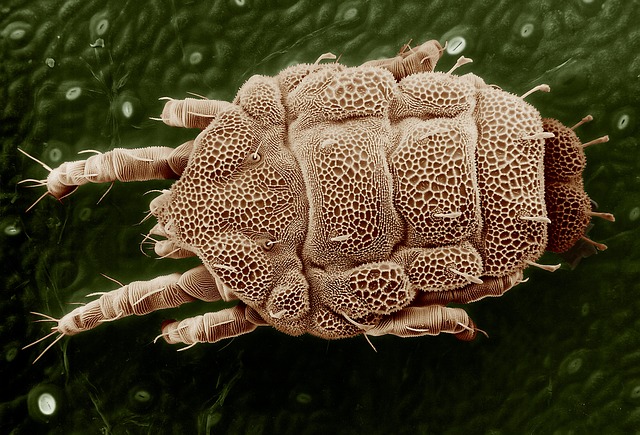
Yellow Mite
All filtrable viruses are obligatory parasites of bacteria, plants, or animals; they have become so biochemically dependent that they can grow and multiply only inside the cells of their hosts. Rickettsias, intermediate between viruses and bacteria, are also obligatory intracellular parasites. Most bacteria are free-living, but many are capable of becoming facultative parasites under favorable conditions. Comparatively few bacteria are true obligatory parasites, causing serious diseases of plants—various blights, wilts, and rots—and many diseases of animals. Yeasts and molds also are mostly free-living; there are among them more parasites of plants than of animals. Some of the fungus plant parasites have extremely complex life cycles involving several hosts.
Every major group of the Protozoa contains parasites. All of the Sporozoa are parasites of vertebrate or invertebrate animals, and the amoebas, flagellates, and ciliates also include parasites of these animals; some flagellates also parasitize plants.
Among the helminths two classes of flatworms, Trematoda and Cestoda (flukes and tapeworms, respectively), are all parasitic in vertebrates. Members of the fluke order Monogenea have simple, one-host life cycles and are ectoparasites on aquatic vertebrates. Flukes of the other main order, Digenea, have complex life cycles involving two or more hosts, the first of which is almost always a mollusk, in which two or more generations are produced by polyembryony, so a single embryo entering a mollusk may produce a million or more larvae. Second intermediate hosts, when utilized, are usually arthropods, fishes, and others that are eaten by the definitive hosts.
Adult tapeworms, which lack an alimentary canal, are parasitic in the digestive tract of vertebrates. Nearly all produce chains of segments, each segment developing complete male and female reproductive systems; this enables them to produce enormous numbers of eggs, up to billions in a lifetime. A few tapeworms also multiply as larvae by budding off several (a great many in the Echinococcus) “heads” or scoleces, each a potential adult. Most intermediate hosts are arthropods or vertebrates eaten by the definitive hosts; some of the Crustacea are probably ancestral hosts.
The spiny-headed worms, Acanthocephala, like the tapeworms, are devoid of an alimentary canal and are parasitic in the digestive tract of vertebrates. They utilize arthropods, usually Crustacea or insects, as first intermediate hosts, and often use second “ecological” hosts to aid them in reaching their definitive hosts.
The nematodes, although the majority are free-living, include many important parasites of both plants and animals. One or another of these may attack any part of a plant or practically any organ or tissue of an animal. Few nematodes reproduce except as mature males or females, but their offspring may number many millions. Some require ecological intermediate hosts, in others these are optional.
Among arthropods four classes contain important groups of parasites—the Crustacea, Arachnida, Pentastomida (Linguatulida), and insects. The Crustacea include many external and a few internal parasites of fishes, and many also attack invertebrates. Nearly all the arachnid parasites belong to the order Acariña, the mites and ticks. All ticks are parasitic and also many mites, though many of the latter are parasitic during only part of their lives.
The Pentastomida are an aberrant group that have become wormlike both in form and life cycle. The adults usually live in the respiratory system of their vertebrate hosts, the larvae in the viscera of some other host.
Among the insects there are parasites in nearly all of the orders, but only lice, fleas, and Strepsiptera are exclusively parasitic. Few except the lice and aphids are parasitic throughout their lives. Lice live generation after generation on one host and are normally transmitted only to mates and offspring, becoming almost a part of the heredity of their hosts, and giving valuable clues to phylogenetic affinities.
Many parasitic insects are such only by virtue of their bloodsucking or sapsucking habits. The sapsuckers are especially numerous in the order Hemiptera (for example, aphids and scale insects), and the bloodsuckers, in addition to lice, fleas, and some bugs (Hemiptera), in the Diptera. Many insects are only parasitic as larvae. Some of these, most of them in the Diptera, attack the tissues of vertebrates, causing a condition known as myiasis. Other larvae, especially in the orders Strepsiptera, Diptera, and Hymenoptera, attack various invertebrates. Most of these eventually completely devour their hosts, thus becoming predators rather than parasites.
Among vertebrates only lampreys and hag-fishes are true parasites in the usual sense of the term. The lampreys, recently established in the American Great Lakes, are now causing enormous loss to the lake fishes.
